



服务热线
178 0020 3020
本期研究
1. 铁催化芳烃C-H羟基化
2. 掺杂莫特绝缘体从极化金属到费米液体的微观演化
3. 利用区域地震网络对灾难性流动事件进行检测和潜在预警
4. 电离液态水中短寿命羟基-水合氢对的成像
5. 通过桥接诱导致密化的高强度可扩展 MXene 薄膜
6. 超顺电弛豫铁电体中的超高能量存储
7. 高度传染性病原体的地方性持续存在:野生动物宿主中的口蹄疫
8. 头颈癌的蛋白质网络图揭示了 PIK3CA 突变的药物敏感性
9. 乳腺癌的蛋白质相互作用图谱
10. 使用蛋白质系统的多尺度图谱解释癌症突变
11. 激活的 IRE1α 的应力感应域在狭窄的 ER 膜管中形成螺旋丝
12. 广泛使用的 IS200/IS605 转座子家族编码多种可编程的 RNA 引导的核酸内切酶
13. 来自芽的光诱导移动因子调节根瘤菌引发的大豆根瘤
14. 通过循环碳经济实现净零温室气体排放塑料

封面:南非克鲁格国家公园的非洲水牛。水牛是口蹄疫 (FMD) 病毒的宿主,可感染野生和家养有蹄类动物。口蹄疫之类的高度传染性感染在易感人群中迅速传播,但随着宿主获得免疫力而逐渐消失——除了在宿主中,它们在地方性流行中持续存在。地方性持续存在的机制代表了长期宿主-病原体关联的进化结果,并可能暴露传播周期中的脆弱点。
8. 头颈癌的蛋白质网络图揭示了 PIK3CA 突变的药物敏感性
A protein network map of head and neck cancer reveals PIK3CA mutant drug sensitivity
我们概述了通过多维蛋白质-蛋白质相互作用图阐明肿瘤遗传复杂性的框架,并将其应用于增强我们对头颈部鳞状细胞癌的理解。该网络揭示了癌症和非癌细胞状态的 771 种相互作用,包括 WT 和突变蛋白亚型。优先考虑富含癌症的相互作用揭示了成纤维细胞生长因子受体酪氨酸激酶 3 与 Daple(一种鸟嘌呤核苷酸交换因子)之间以前未知的关联,导致 Gαi 和 p21 活化蛋白激酶 1/2 的激活以促进癌细胞迁移. 此外,我们观察到人类表皮生长因子受体 3 (HER3) 受体酪氨酸激酶和 PIK3CA(磷脂酰肌醇 3-激酶的 α 催化亚基)之间富含突变的相互作用,可以告知体内对 HER3 抑制的反应。我们预计该框架的应用对于将遗传改变转化为对许多疾病领域潜在生物学的分子和临床理解具有重要意义。

9. 乳腺癌的蛋白质相互作用图谱
A protein interaction landscape of breast cancer
癌症与多种基因组改变有关。为了帮助从机制上理解乳腺癌浸润性癌的这种改变,我们应用亲和纯化-质谱法来描绘 40 种频繁改变的乳腺癌 (BC) 蛋白的综合生物物理相互作用网络,这些蛋白有和没有相关突变,跨越三个人类乳腺细胞系。这些网络确定了癌症特异性蛋白质-蛋白质相互作用 (PPI),它们相互关联并丰富了常见和罕见的癌症突变,这些突变通过引入关键的 BC 突变而大幅重新连接。我们的分析将 BPIFA1 和 SCGB2A1 鉴定为 PIK3CA 相互作用蛋白,它们抑制 PI3K-AKT 信号传导,并发现 USP28 和 UBE2N 作为 BRCA1 的功能相关相互作用物。我们还表明蛋白磷酸酶 1 调节亚基 Spinophilin 与 BRCA1 的去磷酸化相互作用并调节其去磷酸化,以促进 DNA 双链断裂修复。因此,PPI 景观为机械解释疾病基因组数据提供了一个强大的框架,并可以识别有价值的治疗目标。

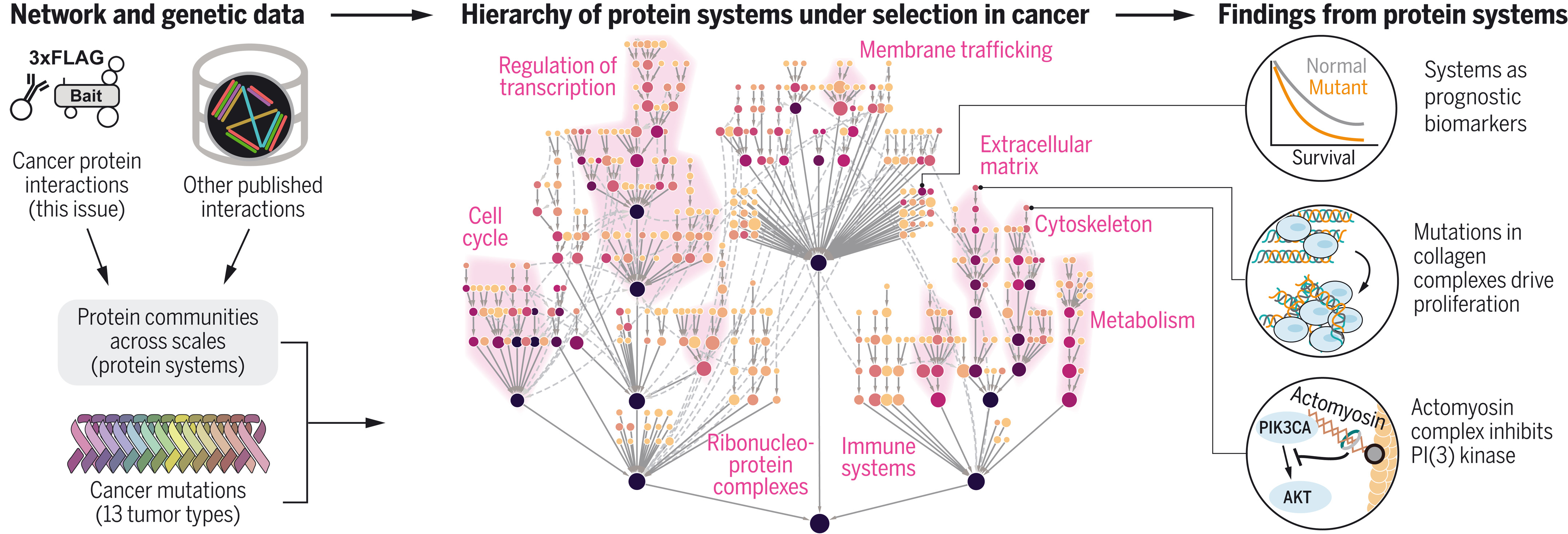
10. 使用蛋白质系统的多尺度图谱解释癌症突变
Interpretation of cancer mutations using a multiscale map of protein systems
癌症研究的一个主要目标是了解分布在不同基因上的突变如何影响常见的细胞系统,包括多蛋白复合物和组装。两个挑战——如何全面地绘制此类系统以及如何识别哪些处于突变选择之下——阻碍了这种理解。因此,我们创建了癌症蛋白质系统的综合图,在多个分析尺度上整合了新的和已发表的多组学相互作用数据。然后,我们开发了一个统一的统计模型,该模型在 13 种癌症类型的突变选择下精确定位了 395 个特定系统。这张名为 NeST(肿瘤嵌套系统)的地图结合了规范过程和显着发现,包括抑制磷脂酰肌醇 3-激酶信号传导的 PIK3CA-肌动球蛋白复合物和促进肿瘤增殖的胶原复合物中的反复突变。这些系统可用作临床生物标志物,并在癌症进化和进展中涉及总共 548 个基因。这项工作显示了不同的肿瘤突变如何在不同尺度上聚合到蛋白质组装上。
11. 激活的 IRE1α 的应力感应域在狭窄的 ER 膜管中形成螺旋丝
The stress-sensing domain of activated IRE1α forms helical filaments in narrow ER membrane tubes
未折叠蛋白反应 (UPR) 的信号网络根据需要调整内质网 (ER) 的蛋白质折叠能力。最保守的 UPR 传感器 IRE1α 跨越内质网膜并通过寡聚化激活。IRE1α 寡聚体在动态病灶中积累。我们通过低温相关光和电子显微镜结合电子冷冻断层扫描和补充免疫电子显微镜在哺乳动物细胞系中确定了 IRE1α 病灶的原位结构。IRE1α 病灶定位于具有复杂分支的狭窄吻合 ER 管(直径,~28 nm)网络。管腔含有蛋白质丝,它可能由排列成两个相互缠绕的左旋螺旋的 IRE1α 腔域二聚体阵列组成。
12. 广泛使用的 IS200/IS605 转座子家族编码多种可编程的 RNA 引导的核酸内切酶
The widespread IS200/IS605 transposon family encodes diverse programmable RNA-guided endonucleases
IscB 蛋白是在不同的 IS200/IS605 转座子家族中编码的推定核酸酶,可能是 RNA 引导的核酸内切酶 Cas9 的祖先,但 IscB 的功能及其与任何 RNA 的相互作用仍未得到表征。利用进化分析、RNA 测序和生化实验,我们从 IS200/IS605 转座子重建了 CRISPR-Cas9 系统的进化。我们发现 IscB 使用单个非编码 RNA 进行双链 DNA 的 RNA 引导切割,并可用于人类细胞中的基因组编辑。我们还展示了 TnpB 的 RNA 引导的核酸酶活性,TnpB 是另一种 IS200/IS605 转座子编码蛋白,也是 Cas12 核酸内切酶的可能祖先。这项工作揭示了一类广泛的转座子编码的 RNA 引导的核酸酶,我们将其命名为 OMEGA(强制移动元件引导的活动)
https://www.science.org/toc/science/374/6563

附件